1 Introduction
The Crow Canyon Archaeological Center launched the Northern Chaco Outliers Project (NCOP) in 2016 (Ryan 2016) to investigate the Lakeview Community, a cluster of four Chaco period great houses in southwestern Colorado. These great houses are located within three archaeological sites: the Haynie site (5MT1905), Ida Jean (5MT4126), and Wallace Ruin (5MT6970). Over the past eight years, most NCOP fieldwork has focused on the Haynie site, a complex ancestral settlement occupied intermittently from approximately A.D. 800 to 1175 (Throgmorton et al. 2022, 2023). The site features two great houses, a large Pueblo I-II roomblock (A.D. 750-1150), and various other structures and contexts throughout the Pueblo I and III periods (A.D. 750-1280).
Analyzing archaeofaunal remains from the Haynie site is a key component of the Northern Chaco Outliers Project. The first year of archaeofaunal analyses revealed a high degree of bone fragmentation and low identifiability, driven by the shaft fragments of medium-to-large mammals (Dombrosky and Gilmore 2023). The preliminary findings also suggested taxonomic diversity estimates were incomplete and animal diversity was not notably high at Haynie compared to selected other sites. Additionally, we identified rare taxa, including wolf (Canis lupus) and bison (Bison bison). The wolf specimen was identified using a highly accurate classification model for canid mandibular measurements. In the second year of faunal analyses (Dombrosky and Feldstein 2024), beyond reporting on a whole new year’s worth of identifications, we summarized the contexts that dogs (genus Canis) have been recovered from in the central Mesa Verde region and how the Haynie site compares. We found no clear associations with typical descriptive categories for animal internments in the U.S. Southwest, and suggest reconceptualization of how dogs fit into categories of deposition, discard, and offering (sensu Monagle and Jones 2020). We also incorporated disturbance contexts into pinpointing the cause of the low faunal identifiability rate at Haynie. We found that disturbance contexts do not strongly influence how identifiable or unidentifiable bone specimens are, further suggesting medium-to-large mammal bone fragmentation could have been driven by the occupants of the Haynie site. Finally, we re-estimated diversity at the site to gauge taxonomic representativeness and compared these estimates to all of the faunal assemblages in the Crow Canyon Research Database. We found—with the addition of new specimens—Haynie had the highest amount of taxonomic diversity compared to other sites in the database, but diversity was not significantly high compared to the other sites.
Here, we report on another year’s worth of faunal identifications, but also provide an in-depth analysis of artiodactyl body size in the central Mesa Verde region (which includes body size estimates from Haynie site specimens). Body size may be influenced by the amount of hunting on large game populations (also known as harvest pressure). This analysis, led by Crow Canyon Zooarchaeology Intern, Chelsea Betts, offers another important way to test for overhunting of large game through time in the central Mesa Verde region (Section 4). We also provide a new analysis associated with Haynie bison specimens. We model the spatial distribution of grass carbon stable isotope (δ13C) values (with what is known as an isoscape) to predict where Haynie bison specimens come from on the landscape. Using this approach, we evaluate whether the Haynie bison specimens were traded in from the Plains or if they come from areas closer to site. Additionally, we compare fragmentation at the Haynie site to all other sites in the Crow Canyon Research Database (Section 5). The larger goal here is to document different research avenues associated with the Haynie archaeofauna and the quality of Haynie site identifications produced from May 2024 through August 2024.
2 Materials and Methods
There are four analysts associated with the data described here: Jonathan Dombrosky, Eric Gilmore, Ahna Feldstein, and Chelsea Betts. Jonathan Dombrosky, Environmental Archaeologist at Crow Canyon, has approximately 12 years of experience with archaeofaunal analysis, and he analyzed new specimens associated with this report from May 2022 to August 2024. Eric Gilmore had approximately three years of experience with archaeofaunal analysis, and he analyzed specimens as a Crow Canyon Zooarchaeology Intern in the summer of 2022. Ahna Feldstein had four years of experience with faunal analysis and was an intern from May 2023 to July 2023. Chelsea Betts had over one year of experience with faunal analysis and was an intern from May 2024 to July 2024. Jonathan, Eric, Ahna, and Chelsea are respectively referred to as Analyst 1, Analyst 2, Analyst 3, and Analyst 4 in all subsequent interanalyst comparisons.
The main comparative collection used for the analysis of the Haynie site archaeofauna is housed in Crow Canyon Archaeological Center’s Laboratory. Three cottontail (Sylvilagus spp.) specimens and one black-tailed jackrabbit (Lepus californicus) specimen were loaned from the Laboratory of Zooarchaeology at the University of North Texas. We took specimens that were difficult-to-identify to the Museum of Southwestern Biology’s Division of Mammals and Division of Birds located in Albuquerque, New Mexico. Various osteological guides, manuals, atlases, and keys aided identification. Several publications assisted with the identification of mammal remains (Adams and Crabtree 2012; Chavez 2008; Gilbert 1980; Hillson 1986, 1996; Jacobson 2003; Olsen 1964; Smart 2009). Bird remains were identified with other works (Cohen and Serjeantson 1996; Gilbert et al. 1981; Hargrave and Emslie 1979; Olsen 1979), as well as fish, amphibian, and reptile bones (Olsen 1964). Some consulted references helped identify both avian and mammalian remains (Broughton and Miller 2016; Elbroch 2006). Yet other works verified nonhuman from human remains (Baker et al. 2005; France 2009; White et al. 2012).
We followed identification protocols explicitly designed to enhance data quality (Driver 1992, 2011; Wolverton 2013; Wolverton and Nagaoka 2018) and used the coding system by Driver (2006). Briefly, analysts adopted a conservative approach to identifying zooarchaeological specimens at the Haynie site. It is an almost impossible task for analysts to understand how all diagnostic skeletal criteria change through time, among species, within different age classes, between sex, and across geographic areas on a fragment-by-fragment basis. It has been argued identifications become less taxonomically specific when analysts have more experience, greater access to diverse comparative materials, and a specific focus on data quality (Gobalet 2001; Lyman 2002, 2019; Wolverton and Nagaoka 2018). This lack of taxonomic specificity likely increases identification accuracy in situations where assemblages contain an abundance of fragmented remains from closely related taxa.
We use the Number of Identified Specimens (NISP) to report taxonomic abundance, and this quantitative unit is a tally of all archaeofaunal specimens within a given taxonomic classification. NISP is the most basic quantitative unit from which most others are derived, such as the Minimum Number of Individuals (MNI). NISP is preferred because it is often highly correlated with measures like MNI. It is also devoid of errors in additive calculation that plague minimum number units (Grayson 1984; Lyman 2008).
All statistical analyses and figures were produced with R version 4.3.3 (R Core Team 2024). Our statistical analyses are structured with tidyverse packages and syntax (Wickham et al. 2019). All graphs were produced with ggplot2 (Wickham 2010).
3 Identified Taxa
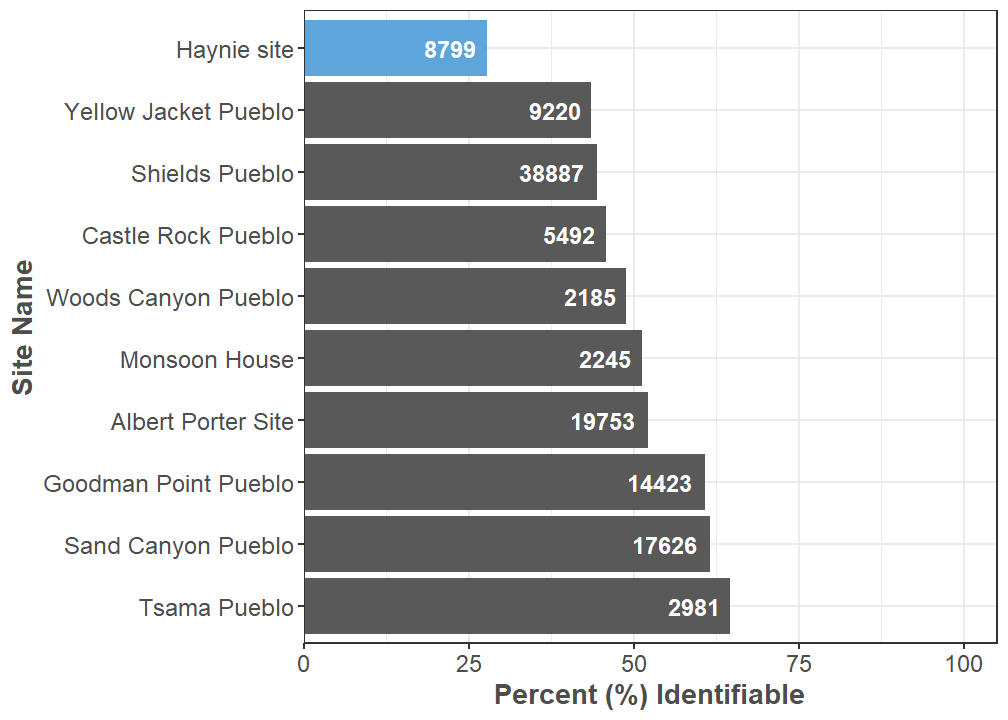
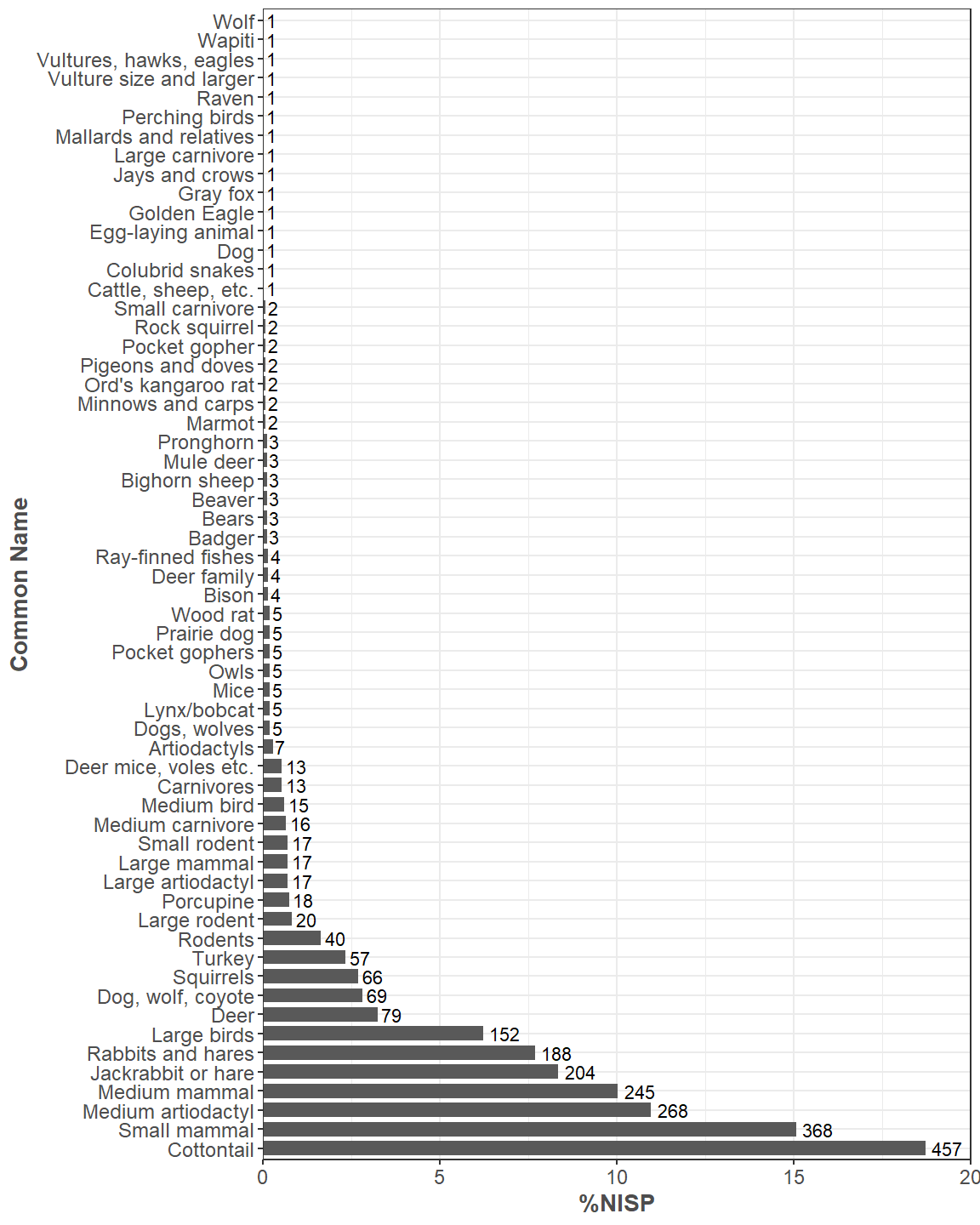
There are 2,440 identifiable and 6,359 unidentifiable specimens to date in the Haynie archaeofaunal assemblage. We have added 249 identifiable and 704 unidentifiable specimens—for a total of 953 specimens—since our last report. Our identification rate is still low at 27.73%, which has mostly gone unchanged. Of all the large faunal assemblages Crow Canyon has analyzed over the years, Haynie currently has the lowest identification rate (Figure 1). Analyst 1 analyzed 60.4% of the assemblage, Analyst 2 analyzed 18.33%, Analyst 3 analyzed 15.07%, and Analyst 4 analyzed 6.19%. There are 4 classes of animals present: Mammalia (mammals), Aves (birds), Actinopterygii (ray-finned fishes), and Reptilia (reptiles). There are 4 orders of mammals present, 7 orders of birds, 1 order of ray-finned fishes, and 1 order of reptile. In total, we used 61 identification types (Figure 2).
3.1 Mammalia (n = 2194)
3.1.1 Lagomorpha (n = 849)
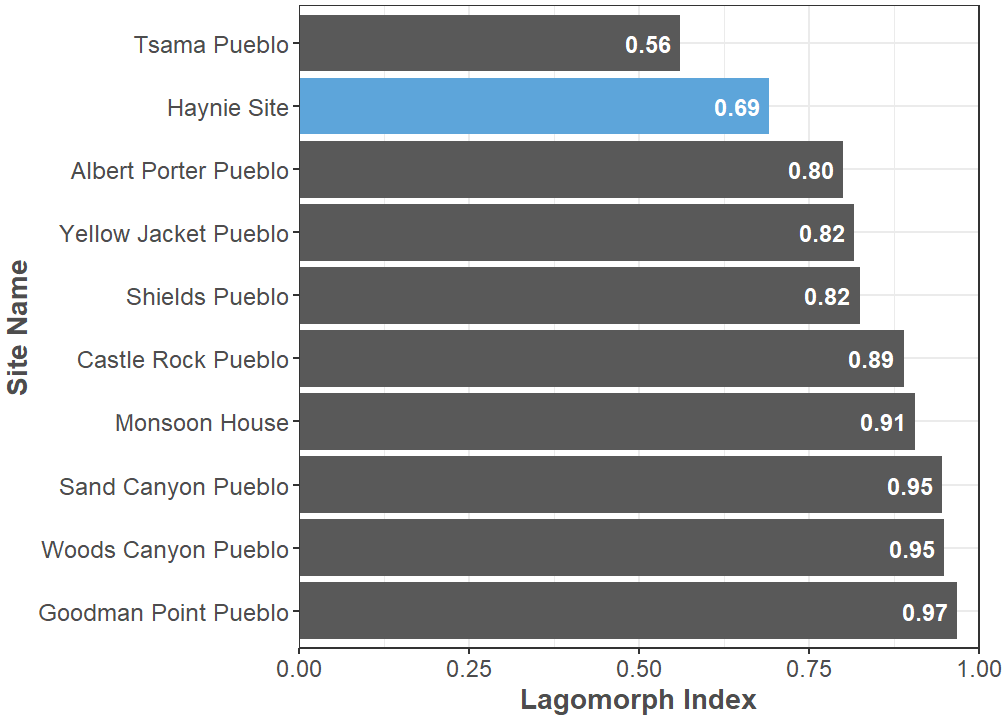
Cottontails (Sylvilagus spp.) and jackrabbits (Lepus spp.) are the main taxa in the order Lagomorpha recovered from the Haynie site. Lagomorphs are currently the most abundant animals identified, comprising 34.8% of the identified specimens. It has been hypothesized populations of larger-bodied jackrabbits decreased through time in the central Mesa Verde region, and this decrease is likely due to human overhunting (Driver 2002; Ellyson 2014). Thus, the ratio of cottontails to jackrabbits—commonly referred to as the Lagomorph Index—is a basic quantitative unit of general interest in the area, and in western North America in general (Driver and Woiderski 2008). The Lagomorph Index at the Haynie site is 0.69, which indicates a fairly equal relationship between the abundance of cottontails and jackrabbits. Haynie currently has the lowest Lagomorph Index of all the large (greater than 1,000 NISP), central Mesa Verde faunal assemblages in the Crow Canyon Research Database (Figure 3). This number is similar to the Lagomorph Index from some Pueblo II (A.D. 950-1150) components of Shields Pueblo (5MT3807). Shields serves as an important point of comparison—here and in subsequent analyses—considering that it and Haynie both have similar site features (i.e., great houses) and general occupation histories (Rawlings 2006).
Garden hunting is an important subsistence practice to consider at the Haynie site, and the moderate Lagomorph Index value is interesting in this regard. Researchers argue that higher ratios of cottontails to jackrabbits indicates a higher reliance on garden hunting in the central Mesa Verde region (Driver 2011). What could the more even relationship of cottontail and jackrabbit abundance indicate about garden hunting at the Haynie site? Future work using stable isotope analysis (sensu Dombrosky et al. 2023) or tooth microwear might help shed light on whether garden hunting was prevalent at the site, and it also might help describe the relationship between the Lagomorph Index and garden hunting in general.
3.1.2 Artiodactyla (n = 390)
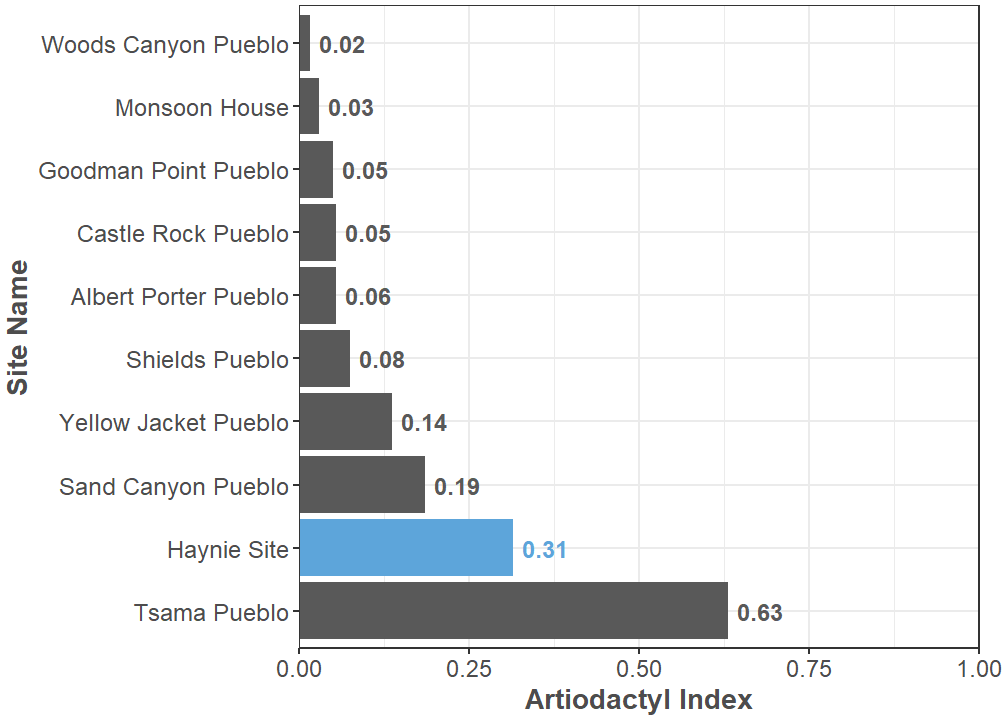
Even-toed hoofed animals make up the order Artiodactyla, and they are currently 15.98% of identified specimens recovered from the Haynie site. Substantial zooarchaeological evidence indicates artiodactyls—mostly mule deer (Odocoileus hemionus), pronghorn (Antilocapra americana), and bighorn sheep (Ovis canadensis)—were severely overhunted in the central Mesa Verde region (Badenhorst and Driver 2009). Some models suggest deer populations might have been so low that any observable deer was immediately hunted after A.D. 1000 (Bocinsky et al. 2012). For this reason, the Artiodactyl Index is another basic quantitative unit of general interest in the region and in the larger northern U.S. Southwest. It measures the ratio of large-bodied artiodactyls to small-bodied lagomorphs in an archaeofaunal assemblage (Broughton et al. 2011). The current Artiodactyl Index at Haynie is 0.31. This value may seem somewhat low, but it is high relative to other sites in the central Mesa Verde region. The Haynie site currently has the highest Artiodactyl Index of all the large (exceeding 1,000 NISP) faunal assemblages in the Crow Canyon Research Database, the only exception is a Pueblo IV (A.D. 1300-1600) site from the Northern Rio Grande called Tsama Pueblo (Figure 4).
Large artiodactyls—elk (Cervus canadensis) and bison (Bison bison)—are also notable parts of the Haynie archaeofaunal assemblage. These specimens comprise 5.64% of artiodactyls. There are currently 4 bison specimens, and Dombrosky and Gilmore (2023) reported on these specimen identifications in detail. Importantly, these specimens do not overlap with each other and represent an MNI of 1.
Dombrosky and Feldstein (2024) reported on radiocarbon dates from the bison specimens to potentially gauge if two or more individual bison were present at the Haynie site. There was almost complete overlap in the 95% calibration date ranges, and two specimens had the same exact conventional radiocarbon age estimation. These results suggest that it is possible all the specimens belong to the same individual. However, given that the calibrated date ranges are around 150 years, it is still possible they could be from two or more individuals. Regardless of how many individuals were present at Haynie, there is another interesting question surrounding these specimens: where did the bison individuals actually come from?
The procurement area for bison is a classic topic of research in Southwest zooarchaeology. The typical question is: were bison hunted locally or were they part of exchange relationships with more mobile Plains groups (sensu Speth 1991)? The central Mesa Verde region does not come to mind as typical habitat for massive, roving herds of bison. However, it has been argued that small bands of these animals migrated through the Colorado Plateau since the late Pleistocene and into the Holocene (Martin et al. 2017). Thus, that bison were procured somewhat closer to the Haynie site than from the plains is a valid question in need of testing. Here, we use stable isotope values to predict where on the landscape Haynie bison specimens fed from before their deaths. We use δ13C isoscape models of grasses for this task. Specifically, we rely on a widely used modern isoscape model provided by Cotton et al. (2016) and constructed using bison bone collagen and tooth enamel sampled from museum collections (Figure 5). We also used the raw data from Cotton et al. (2016) and created a new isoscape model that takes elevation into account using the IsoriX package. This newer package has the added benefit of providing statistical predictions for where on the landscape unknown samples derive from.
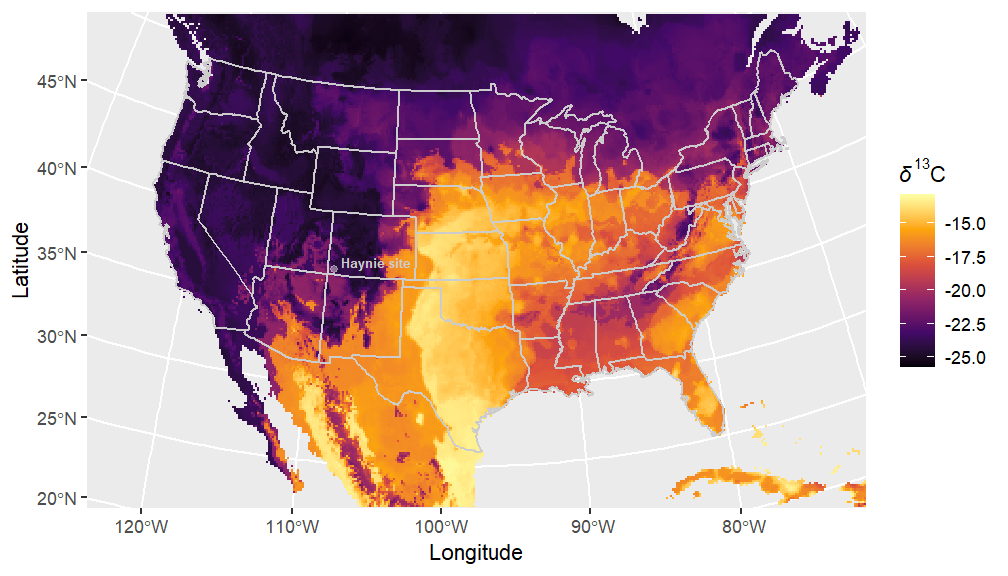
The five Haynie bison bone collagen specimens’ have δ13C values ranging from -24.2‰ to -23.0‰. These values have the same Trophic Discrimination Factor (-4.875‰) incorporated that Cotton et al. (2016) used. These δ13C values do not occur in the Plains, but are represented in the general Four Corners region where the Haynie site is located (Figure 6). The new isoscape model developed here (Figure 7) also supports this interpretation (Figure 7). It is important to note these models do not provide fine scale resolution for where exactly these specimens occur on the landscape, but it is possible the Haynie specimens could come from higher elevation areas like the La Plata Mountains to the east (Figure 8, Figure 9).


IsoriX package. The main difference is that the IsoriX model incorporates elevation into the isoscape modeling algorithm. The location of the Haynie site is marked by the white circle.
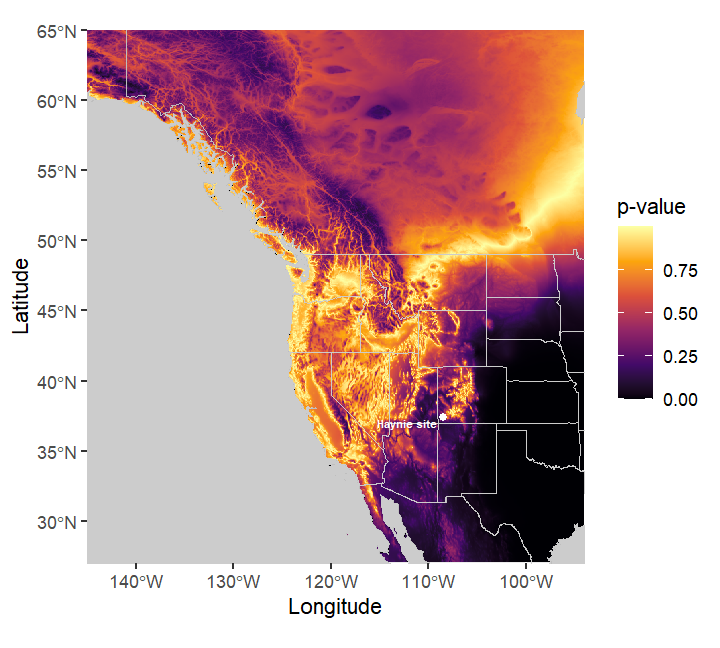
IsoriX isoscape model to predict where Haynie site (marked by the white circle) bison specimens could likely come from. The prediction model here tests where specimens do not come from. Thus, lower p-values (darker areas) indicate where specimens most likely do not derive from. The individual (or individuals) that comprise the Haynie specimens likely did not come from the plains.
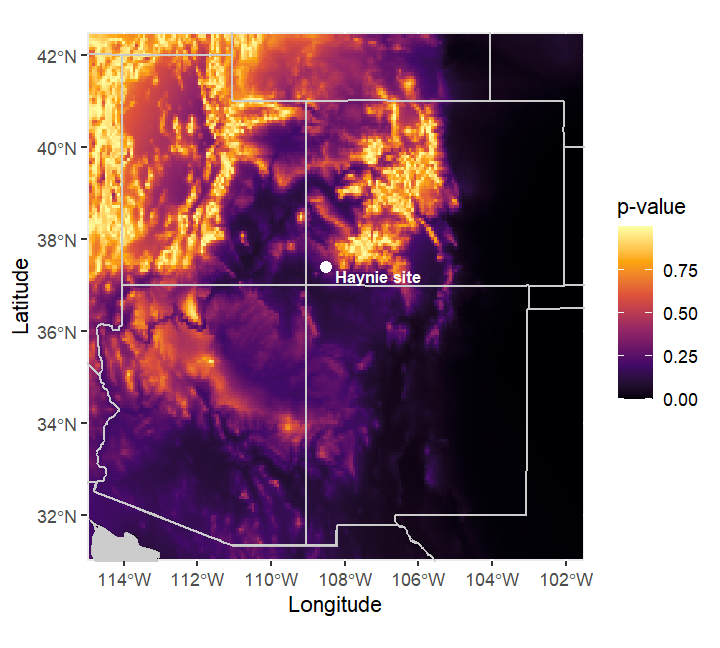
IsoriX prediction model (Figure 8) to show areas around the Four Corners region where the bison specimens could derive from.
3.1.3 Small Mammal (n = 368)
This identification group includes those mammals jackrabbit size and smaller. This non-standard identification includes all small mammal specimens lacking morphological features required for more specific taxonomic levels. Consequentially, it has a high likelihood of incorporating many lagomorph specimens since they are exceedingly abundant in southwestern archaeofaunas. This identification group will be incorporated into final comparisons of size-based abundance indices through time, between areas of the Haynie site, or between sites. The goal will be to gauge the impact this identification group has on final interpretations of the Lagomorph Index.
3.1.4 Medium Mammal (n = 245)
Mammals larger than a jackrabbit and up to deer size are considered medium mammals. There is a high probability this identification group incorporates many artiodactyl specimens since they are one of the most common medium mammals in southwestern archaeofaunas. It should be incorporated in sensitivity analyses that rigorously assess final conclusions relying on size-based abundance index values, such as the Artiodactyl Index.
3.1.5 Rodentia (n = 205)
Rodents are 8.4% of identified specimens, which is a small component of the overall assemblage. The ratio of sciurids to rodents can help gauge the impact of intrusive species. Members of the squirrel family (Sciuridae) are notorious intruders of archaeological deposits, especially prairie dogs (Cynomys spp.). The majority of prairie dog skeletal fragments are identified to the family level, as it is extremely difficult to skeletally distinguish prairie dogs from ground squirrels. This means Sciuridae identifications have the highest potential for accumulating prairie dog specimens. Indeed, we confirmed that the vast majority of sciurid specimens compare favorably to prairie dogs. Sciurids make up 36.59% (n = 75) of the rodent assemblage. Given that rodents comprise a small portion of the overall assemblage, it does not appear that rodent intrusions pose a significant problem for interpretation at the site. We will, however, closely track the taxonomic composition of rodents as the Northern Chaco Outliers Project progresses. Importantly, rodents could have been actively hunted (Badenhorst et al. 2023). One way to disentangle intrusive from non-intrusive rodents is through radiocarbon dating (Guiry et al. 2021). This could prove useful if prairie dog specimens preclude clear interpretation of the Haynie archaeofaunal assemblage in the future.
Large rodents, those larger than a woodrat (Neotoma spp.), comprise 20.98% of the total rodents currently identified. Beaver (Castor canadensis) and porcupine (Erethizon dorsatum) specimens are the most notable. Beavers prefer aquatic habitats while porcupines take refuge in trees along active floodplains (Baker and Hill 2003; Roze and Ilse 2003). The presence of these species might suggest hunting activities focused in riparian habitat close to the site.
3.1.6 Carnivora (n = 120)
Carnivores are a small portion of currently identified specimens at the Haynie site (4.92%), but a number of canid specimens were discussed in detail in previous reports (Dombrosky and Gilmore 2023; Dombrosky and Feldstein 2024). There are currently 77 specimens from the family Canidae in the Haynie site fauna. Many of the specimens come from an articulated domestic dog offering and one specimen is a wolf mandible.
3.1.7 Large Mammal (n = 17)
The large mammal identification includes mammals larger than deer, and it includes specimens lacking morphological features required for more specific taxonomic levels. It is likely that it could incorporate large artiodactyls like elk and bison specimens. It is recommended it be incorporated in sensitivity analyses that rigorously assess conclusions based on these animals.
3.2 Aves (n = 238)
3.2.1 Large Birds (n = 152)
Birds larger than a mallard are considered large birds. This identification group is most likely dominated by Turkeys (Meleagris gallopavo) as they are one of the most frequent birds recovered from ancestral sites in the region. A high proportion of Turkey specimens are assigned to this group considering there is considerable skeletal morphological overlap between Sandhill Crane (Grus canadensis) and Turkey (Hargrave and Emslie 1979). The Crow Canyon comparative collection does not, as of yet, include Sandhill Crane skeletal material. As a result, this group is frequently used as it represents a broader identification category.
3.2.2 Galliformes (n = 57)
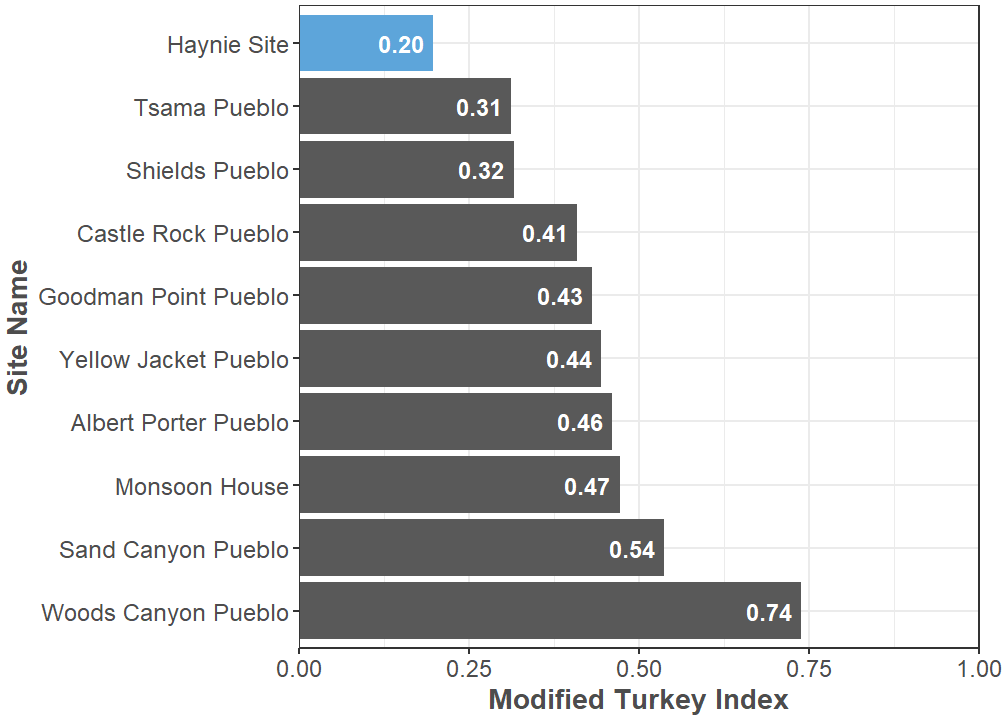
All Galliformes specimens identified so far in the Haynie assemblage are Turkey, but it is still surprising how few Turkey specimens there seem to be. For instance, Spielmann and Angstadt-Leto (1996) proposed the Turkey Index, which is the ratio of Turkeys to lagomorphs in an assemblage. The current Turkey Index for Haynie is 0.07, which is notably low. However, Driver (2002) proposed the Modified Turkey Index where the large bird identification group is included in the calculation. The current Modified Turkey Index value for Haynie is 0.20. This value is within the normal range for Pueblo I/Pueblo II period sites in the central Mesa Verde region (Badenhorst and Driver 2009), but it is the lowest Modified Turkey Index for large assemblages in the Crow Canyon Research Database (Figure 10). Also worth noting, there appears to be an abundance of turkey specimens yet to be identified. It is doubtful that this low index value will remain constant as the faunal sample increases. How turkey husbandry was managed at the community-level is an essential future area of research for the Northern Chaco Outliers Project. This line of inquiry is one way to delve deeper into aspects of cooperation and identity at the Lakeview Community.
3.2.3 Medium Birds (n = 15)
Medium birds are considered larger than a robin and the size of a mallard or smaller. It is difficult to attribute the majority of this identification group to a single taxon, as it contains a variety of difficult-to-identify fragmented skeletal parts that could belong to numerous taxa.
3.2.4 Strigiformes (n = 5)
The owl specimens identified at the Haynie site are a terminal phalanx, second phalanx, first phalanx, femur, and ulna. Owl feathers are known to have been incorporated into dance paraphernalia and prayer sticks (Ladd 1963). It is, however, important to keep in mind that owls can be active taphonomic agents. Luckily, their signatures are well-known and include the presence of pellets, small mammal remains with little-to-no fragmentation, and visible signs of digestion on specimens (Andrews and Cook 1990; Fernández-Jalvo and Andrews 2016). Owls do not appear to be a taphonomic agent of concern at Haynie (Section 5).
3.2.5 Accipitriformes (n = 3)
Accipitriforms are birds of prey active during the day, which include hawks, eagles, vultures, and kites. Two specimens were identified as general members of this order: one general foot phalanx and one terminal phalanx that compared favorably to a Turkey Vulture (Cathartes aura). The other specimen was a distal tibiotarsus fragment identified to Golden Eagle (Aquila chrysaetos). This specimen was taken to the Museum of Southwestern Biology’s Division of Birds and identified with their skeletal comparative collection. It was distinguished from Bald Eagle (Haliaeetus leucocephalus) based off the morphology of the supratendinal bridge, which is more convex and more proximally robust in Golden Eagles compared to Bald Eagles. Eagles are acutely significant in Pueblo culture and their feathers are highly valued (Beaglehole 1936; LaZar and Dombrosky 2022; Tyler 1991).
3.2.6 Passeriformes (n = 3)
Three passeriform (i.e., song bird) specimens have been identified thus far in the Haynie assemblage: one small fragmented humerus generally identified to the order-level, a tibia fragment identified to Corvidae (the family comprising jays and crows), and one large carpometacarpus identified as Raven (Corvus corax).
3.2.7 Columbiformes (n = 2)
Pigeons and doves are in the order Columbiformes, and one ulna fragment and one sternum fragment were recovered from Haynie. These specimens are most likely a Mourning Dove (Zenaida macroura). An order-level identification was used because the Crow Canyon comparative collection does not include a Band-tailed Pigeon (Columba fasciata), which is the only other member in the order Columbiformes to consider in the region.
3.2.8 Anseriformes (n = 1)
One complete carpometacarpus was identified to the genus Anas spp. (mallards and other relatives) from the Haynie site. While this specimen could likely represent the common mallard (Anas platyrhynchos), Hargrave (1965) mentions the likely presence of Mexican Duck (Anas diazi) in the Four Corners region in the past. He based this assertion on the recovery of Mexican Duck feathers associated with Tusayan Polychrome sherds from Glen Canyon. The presence of different mallards and ducks is of potential biogeographic significance and the different members of this genus represent difficult-to-separate taxa.
3.3 Actinopterygii (n = 6)
This taxonomic class includes the ray-finned fishes. Six specimens have been identified to this general class: two ribs and three fragmented vertebrae centra (of which two of these specimens can be identified to the order-level, see below). Following Nelson (2006, 35), fish specimens should no longer be referred to as pisces, as it is an antiquated taxonomic term. Similarly, for fishes of inland North America, the use of osteichthyes should no longer be used (Nelson 2006, 83). This is so for two interrelated reasons. First, this term has been replaced by the Euteleostoma designation. It successfully describes a monophyletic clade that includes tetrapods. Secondly, since Euteloestoma includes tetrapods, it includes lobe-finned fishes (Sarcopterygii). Lobe-finned fishes—like the coelacanth (Actinistia)—are not native fishes in inland North America during the late Holocene (Cloutier and Forey 1991). The use of osteichthyes should be accordingly abandoned. Instead, Actinopterygii (ray-finned fishes) should be used because it is a more accurate class-level designation for archaeofaunas from the U.S. Southwest/Mexican Northwest.
3.3.1 Cypriniformes (n = 2)
This order includes carps, minnows, and suckers, which are common fishes in the aridland streams of the U.S. Southwest (Minckley and Marsh 2009; Sublette et al. 1990). These two specimens are small intact vertebrae. The lateral ridge morphology of centra can be used to identify vertebrae of fishes from U.S. Southwestern archaeofaunas to the order-level. Common orders of fishes found in rivers in the U.S. Southwest include Cypriniformes, Siluriformes, Lepisosteiformes, and Salmoniformes, and each of these orders have distinct vertebral morphology. These specimens are also notably small. It is possible inhabitants of the Haynie site used non-targeted methods to capture fishes, such as seining (Dombrosky et al. 2022). A focus on fishing practices offers a basic way to understand aquatic habitat use associated with the Simon Draw watershed.
3.4 Reptilia (n = 1)
3.4.1 Squamata (n = 1)
One colubrid (family Colubridae) vertebra was identified from Haynie. This specimen was identified to the family-level using Olsen (1964). It likely represents a bullsnake (Pituophis catenifer sayi), which is one of the most common colubrids in Cortez, CO. This taxon frequently overwinters in covered archaeological deposits in the Mesa Verde region and could represent intrusive specimens.
3.5 Oviparous Animals (n = 1)
Eggshell specimens were identified as oviparous animals. These specimens are likely from turkey, but a general identification was given because a scanning electron microscope could not be used to assess mammillary cone morphology (Beacham and Durand 2007; Conrad et al. 2016; Lapham et al. 2016). It is possible, though unlikely, that this eggshell could be from a lizard.
4 Mule Deer Body Size Through Time in the Central Mesa Verde Region
One of the largest zooarchaeological questions in the central Mesa Verde region centers on overhunting and resource depletion. The migration of ancestral communities out of the region—during the end of the A.D. 1200s—is thought to be at least partially caused by a reduction in large game (Muir and Driver 2002; Schollmeyer and Driver 2023). The increased use of turkeys and other smaller game (i.e., cottontails and jackrabbits) is also linked to large game resource depression (Muir and Driver 2002; Badenhorst and Driver 2009; Munro 2011; Ellyson et al. 1990; Schollmeyer and Driver 2023). Artiodactyls, such as mule deer, are the preferred taxa of hunters in the Southwest because of their high caloric return and perhaps even their association with prestige (Schollmeyer and Driver 2023). Overhunting is the current leading explanation of the lack of mule deer in Pueblo III period (A.D. 1150-1280) archaeofaunal assemblages (Muir and Driver 2002; Badenhorst and Driver 2009; Schollmeyer and Driver 2023). However, this interpretation is based on relative abundance data and abundance indices (i.e. the artiodactyl index), which are derived from site-level bone counts (specifically quantified using the Number of Identified Specimens or NISP). While crucial for initial estimations of taxonomic abundance, accurate bone counts are influenced by a multitude of factors. Environmental shifts, human butchery practices, post-depositional processes, overhunting, and many other processes can significantly alter accurate estimations of large game from archaeofaunal assemblages (Lyman 1994). The extent to which each of these processes affected assemblages cannot be confirmed without additional complimentary evidence.
Overhunting remains a hypothesis in the late prehispanic central Mesa Verde region, though multiple strong case studies suggest it occurred (Muir and Driver 2002; Badenhorst and Driver 2009; Schollmeyer and Driver 2023). Multiple lines of evidence, beyond bone counts, should be used to further evaluate this hypothesis. If overhunting is truly the cause of the low relative abundance of mule deer, its signature should also be visible in body size changes and mortality profiles.
The overhunting hypothesis should be rigorously evaluated because the implication that Ancestral peoples were unsustainably using resources is serious. Interpretations like this affect the public perception of Ancestral groups, and as a result, their descendants. One can also extrapolate this hypothesis to imply, on a larger scale, that humans in general cannot avoid unsustainable environmental practices. This section strives to address the question of overhunting through changes in mule deer body size and mortality data, which adds to previous studies and can hopefully create a clearer picture of hunting practices through time and at the end of the Pueblo III period.
4.1 Large Game Body Size and Overhunting
Zooarchaeologists commonly estimate animal body size from archaeofaunal specimens to understand past human hunting strategies, as well as other environmental or climatic factors (Broughton et al. 2011; Dombrosky et al. 2022; Purdue 1987, 1989; Wolverton 2008; Wolverton et al. 2007, 2008, 2009). Deer body size typically tracks with the amount of available food resources in the environment, which is controlled by both anthropogenic and non-anthropogenic factors. Hunting pressure is one anthropogenic factor of particular interest, and this term can be used interchangeably with harvest pressure. Hunting pressure is defined as the degree to which a prey population is thinned by hunting activities (Wolverton 2008). Changes in the degree of harvest pressure can cause changes in food availability and thus body size.
Under the influence of high human hunting pressure, there are fewer individual deer present on the landscape. This results in less competition for food resources among deer. In turn, increased access to food resources translates to the increased body size of individual deer in the population. The population of deer in this scenario is maintained below ecological carrying capacity, which is the total number of organisms that can be supported by an ecosystem over time (Wolverton 2008). The environment can adequately support this population of deer under heavy harvest pressure (and the other organisms in the ecosystem) without strain, which keeps the deer population in a constant state of growth through successive generations (Wolverton 2008; Wolverton et al. 2007, 2008, 2009). Deer populations are higher under the influence of low hunting pressure due to decreased mortality. In this scenario, we would expect to see more individuals present and competing for resources on the landscape. Each individual animal would have fewer available food resources because the population is pushed above or nearer to carrying capacity. As a result, these deer would obtain a smaller body size through time.
Much research on body size has been done on white-tailed deer specifically because of a trait known as phenotypic plasticity. Phenotypic plasticity is the influence of environmental changes and food availability causing change in an observable trait, such as growth rate (Wolverton 2008). The body sizes of white-tailed deer are sensitive to changes in resource availability, which means the maximum body size of these animals can change within just one generation. Mule deer, which are in the same genus as white-tailed deer, also exhibit variability in this trait. However, mule deer body size does not change as dramatically in relation to food availability because these animals are adapted to tolerate a somewhat less nutritious diet (Staudenmaier et al. 2021). While differences in phenotypic plasticity exist between these two species, we argue that the life history strategies of white-tailed deer represent an accurate general analog to mule deer in the case of the late prehispanic central Mesa Verde region. It is reasonable to expect that changes in harvest pressure and food availability would influence mule deer body size and that such changes should be observable in the paleozoological record.
4.2 Body Size Estimation Methods
To estimate large game body size, Analyst 4 measured 35 medium artiodactyl astragali following Purdue (1989) and Wolverton et al. (2007) (Figure 11). Astragali have been commonly used in past research to estimate body size because they are load-bearing skeletal elements. Their size and shape changes based on the locomotion of the animal. They are also very dense, thus they tend to survive in archaeological contexts.
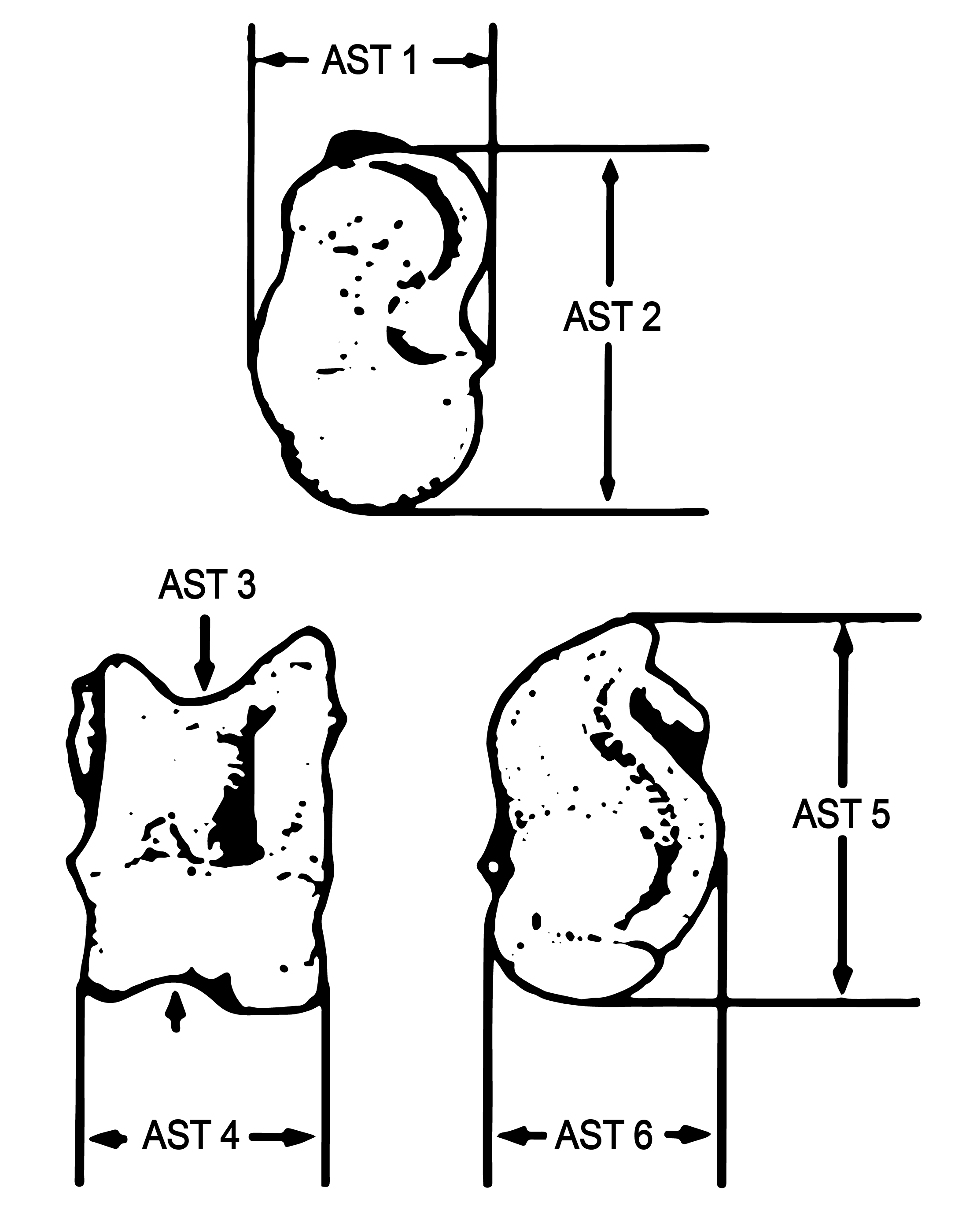
Our astragali assemblage is comprised of several different sites excavated by Crow Canyon Archaeological Center that span a wide range of time periods. All of these astragali are curated by Canyons of the Ancients Visitor Center and Museum. The astragali were recovered from six different sites: The Dillard site (5MT10647; A.D. 420–725), the Haynie site (5MT1905; A.D. 950-1150), Albert Porter Pueblo (5MT123; A.D. 1050–1300), Yellow Jacket Pueblo (5MT5; A.D. 1050–1300), Sand Canyon Pueblo site (5MT765; A.D. 1150–1300), and Goodman Point Pueblo (5MT604; A.D. 1260–1275).
To understand the change in mule deer body size, we first differentiated between artiodactyl species to confirm specimens were not actually white-tailed deer. The Medium Artiodactyl designation follows Driver’s (2006) CCAC manual and is used when a genus or species cannot be identified using distinct morphological criteria. To simplify this task, Analyst 4 re-identified all astragali based on the CCAC comparative collection along with Lawrence’s (1951) criteria and personal notes by other zooarchaeologists. The goal was to limit inter-analyst variability and sort out possible bighorn sheep and pronghorn antelope. To distinguish mule deer and white-tailed deer, Analyst 4 measured all astragali following Dombrosky and Gilmore (2023) (Fig. 2). We built models using modern white-tailed deer and mule deer astragali with known identifications. We used Linear Discriminant Analysis (LDA) as an engine to distinguish archaeological white-tailed deer and mule deer. Our model confidently predicted the assemblage only included mule deer. The models were 98–100% accurate in discriminating between the two species.

4.3 Results
One of the most interesting results of our body size measurements is the wide variation in astragali size towards the end of the Pueblo III period, especially at Sand Canyon Pueblo. We expect mule deer to have a larger body size at the end of Pueblo III if harvest pressure had increased to a point where deer populations were pushed below environmental carrying capacity. However, there is a highly diverse range of astragali size present here, which does not provide resolution on the impacts of harvest pressure. To explore our dataset, we fitted a linear model, a curvilinear model, and a multi-curvilinear model (Figure 13) to our astragali measurements. These are three possible interpretations of mule deer body size through time in the central Mesa Verde region.
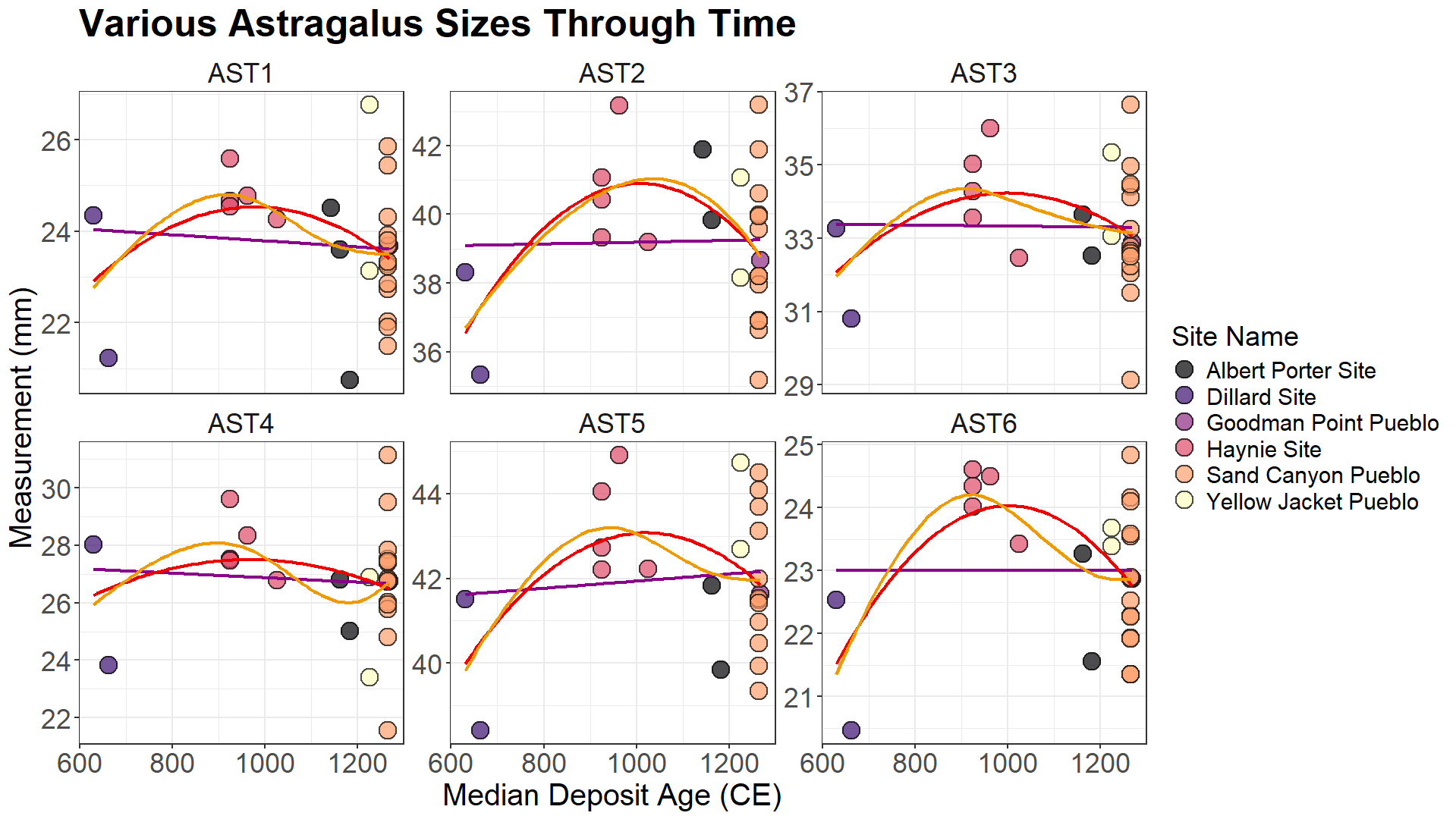
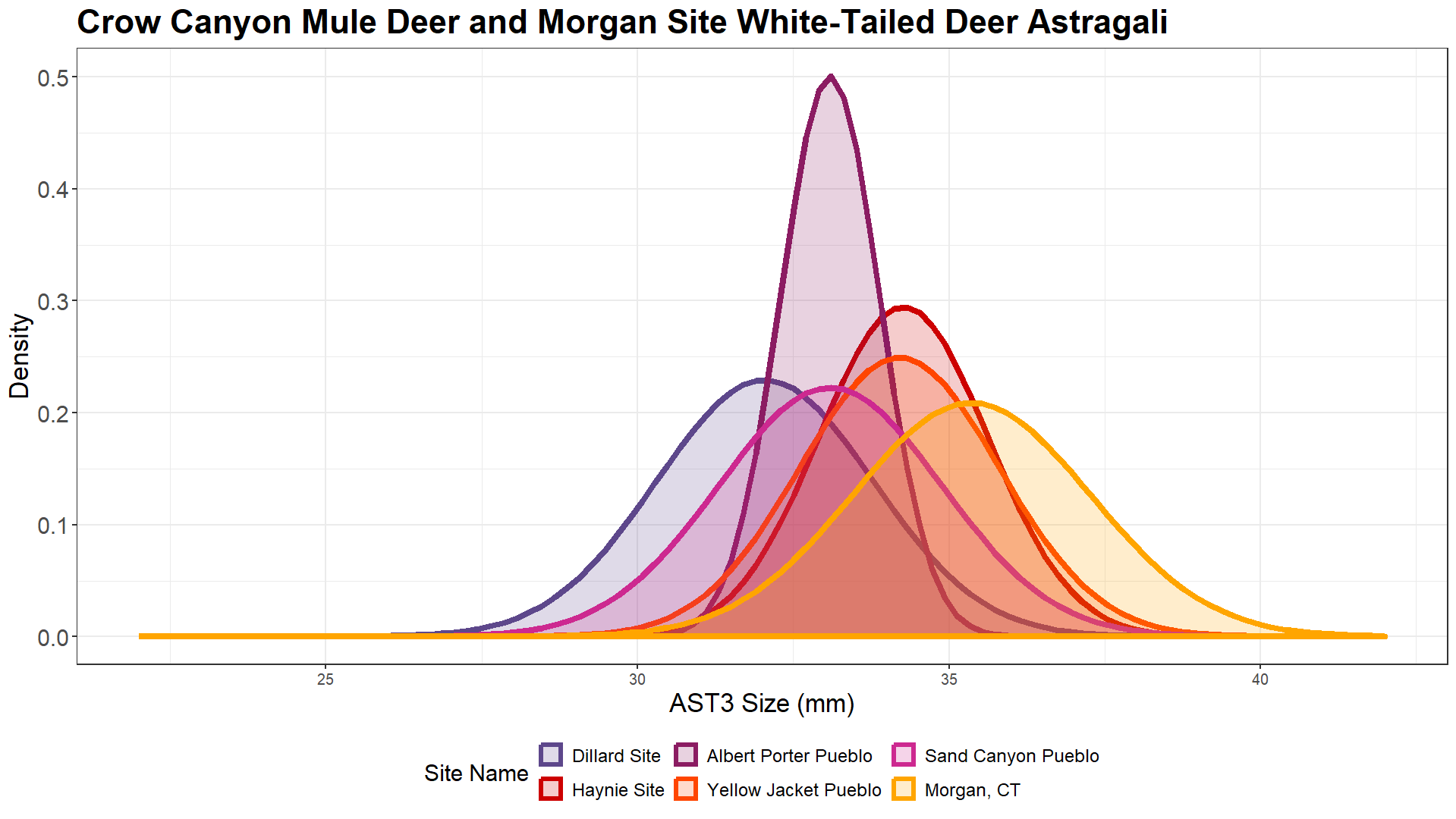
In order to contextualize our data, we compared them to archaeological white-tailed deer assemblages from Weitzel (2023) (Figure 14 and Figure 15), and specifically, the Morgan Site. The Morgan Site is a Late Woodland Period site in Connecticut, and it provides an example of sustainable harvest pressure. The white-tailed deer at the site have a large body size because their habitat was anthropogenically expanded. The environmental carrying capacity increased through agricultural land-clearing and controlled burns, causing deer body sizes to grow larger without extreme harvest pressure (Weitzel 2023). Deer generally prefer to live on the edges of the forest, near disturbed land, thus these changes to the landscape increased their access to resources.

Mule deer body sizes at the Crow Canyon sites overlap with, and fall slightly below, the Morgan Site white-tailed deer body sizes (Figure 15). There are a few ways to interpret this pattern. One possibility is body size did not increase around the end of Pueblo III period. The white-tailed deer body size at the Morgan Site roughly plots around where mule deer might plot if the body sizes had increased. Mule deer body sizes tend to be larger than white-tailed deer body sizes, with significant overlap in their size ranges (Staudenmaier et al. 2021). Another interpretation is because there is a significant amount of overlap between mule deer and white-tailed deer astragali sizes here, the size of mule deer at the central Mesa Verde region sites are actually indicative of healthy and resilient deer populations. Yet another possibility is that high ecological productivity of the environment surrounding the Morgan site is causing white-tailed deer to surpass mule deer in body size. Ecological productivity can improve ontogenetic growth rate and increase maximum body size.
Another important line of evidence to consider is the age at which animals died, which is estimated using morality profiles. Mortality profiles use epiphyseal fusion data to represent the age class of animals at the time of their death. They can be used as an additional method to tease apart factors affecting body size change. Environmental changes and harvest pressure have distinct signatures with some overlap (Wolverton 2008; Wolverton et al. 2008). High harvest pressure (and overhunting) is indicated by an inordinate number of juveniles in the archaeological record. This assumes that energy maximization logic of optimal foraging theory holds true, which postulates the largest individuals with the most amount of meat (and thus energy potential) will be preferentially hunted. Young individuals would have less of an opportunity to get old as the number of larger adult animals dwindle. Low harvest pressure is indicated by a larger presence of older adults in an assemblage. Young individuals would have the chance to grow to older age classes before they are hunted. If there is a clear change in body size, but all age classes in the mortality profile are affected equally, including reduced fertility, that is an indicator of a change in environmental carrying capacity. The quality and quantity of resources available to mothers affect their ability to have young.
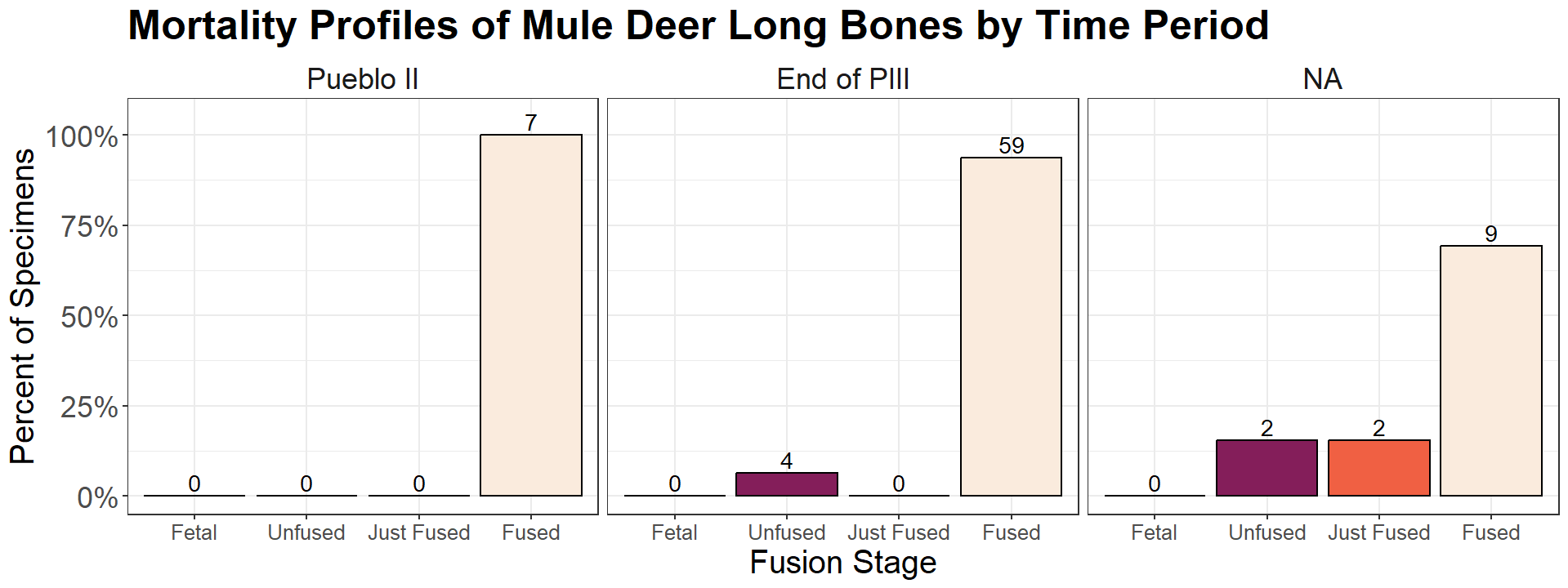
The substantial number of fused long bones at the end of PIII is a sign harvest pressure in the region may not have been as high as previously thought (Figure 16). Mule deer are seemingly given the chance to reach adulthood before they are hunted. Optimal foraging theory is not the only subsistence model possible, but the purpose of its use here is to question the assumption that overhunting occurred at the end of Pueblo III period. Our models do not provide exact snapshots for what harvest pressure and environmental conditions looked like at the end of Pueblo III. They are simply meant to call for the use of multiple lines of evidence in understanding the nuances of harvest pressure, especially at such a pivotal time in human history.
4.4 Conclusions
Preliminary investigations of several new lines of evidence concerning mule deer at the end of Pueblo III period are inconclusive, but they suggest areas for further consideration and study. The diversity of astragali sizes at PIII sites such as Sand Canyon Pueblo shows there are likely factors outside of harvest pressure at play. Body size does not clearly increase as expected under heavy harvest pressure. The body size of white-tailed deer at the Morgan Site overlaps with and/or surpasses the size of mule deer from the central Mesa Verde region. The Morgan Site is an interesting comparison considering it is located in central Connecticut and argued to have unusually large white-tailed deer that were sustainably hunted. The mortality profiles created here show an abundance of adult individuals at the end of PIII, which is another sign that high harvest pressure may not be the main actor on these assemblages.
Regardless of whether there was truly low or high artiodactyl harvest pressure during the late PIII period in the central Mesa Verde region, our point here is simple: the archaeological picture may not be as clear cut as previously thought. More data must be obtained to accurately identify causes of the patterns found here, as sample size is concern in the above analysis. In particular, more astragali measurements are needed from PI and PII periods. More accurate comparisons for archaeological mule deer data should also be developed using modern or archaeological mule deer astragali data with known context and environmental conditions. Future research should be done to create parameters around the sensitivity of mule deer body size to harvest pressure. Most body size models have been created using white-tailed deer. Considering the differences between the diets and morphology of white-tailed deer and mule deer, variation in phenotypic plasticity between these two species should be gauged. Another important thing to understand is how slight differences in the environments surrounding our selected sites can affect body size independently of harvest pressure. Additionally, the consideration of other models beyond optimal foraging theory is also one avenue for future research.
5 Taphonomy
One of the primary taphonomic questions surrounding the Haynie site archaeofauna is: what is causing the large amount of unidentifiable specimens at the site? The last two reports have focused on building predictive models to investigate what taphonomic variables are most associated with unidentifiable specimens. In year one, we found unidentifiable specimens are usually shaft fragments from medium-to-large mammals. In year two, we demonstrated there was an association between unidentifiability and recently disturbed contexts at Haynie, but that this association was of little practical significance (due to low effect size values). Our current hypothesis is medium-to-large mammal processing from Haynie’s Ancestral occupants is the primary cause of unidentifiability and recent archaeological deposit disturbance is slightly exaggerating the signal of unidentifiability.
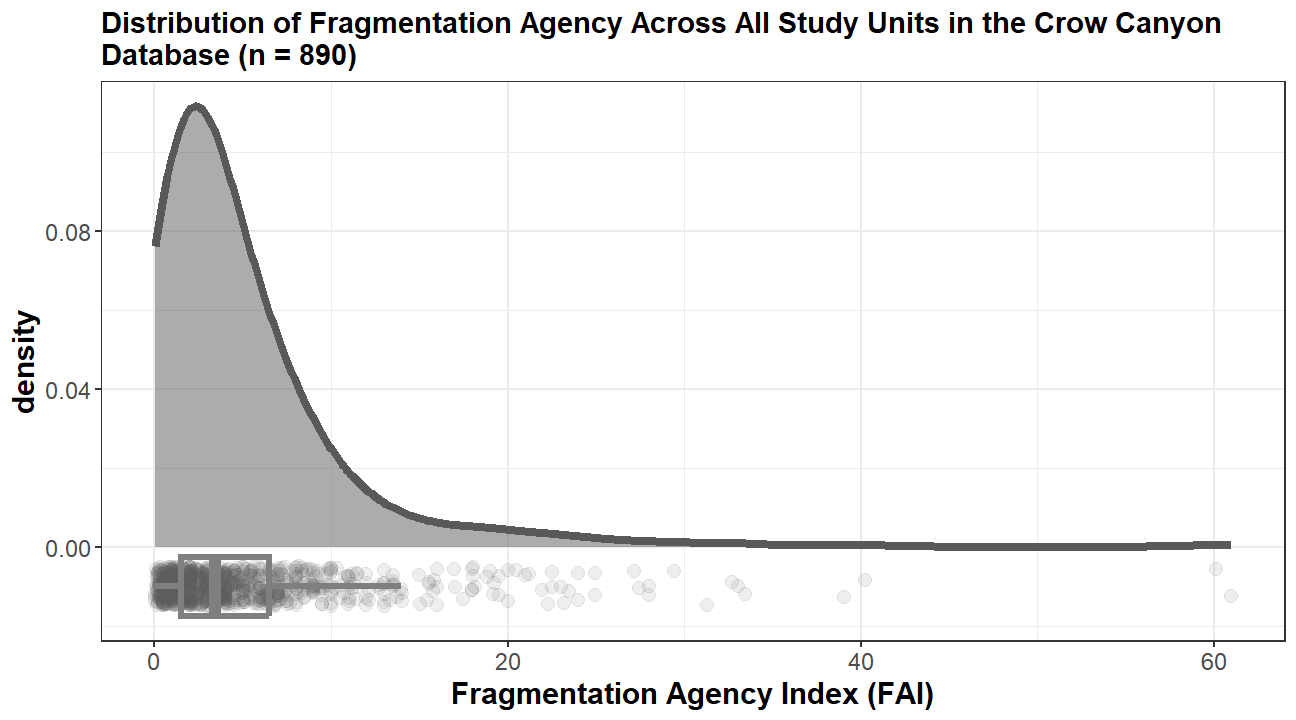
In this report, we step back from predictive modeling, zoom out, and spend time thinking about Haynie from a comparative taphonomic perspective. This last year, we worked to better understand taphonomic variables across the Crow Canyon Research Database. In particular, we focused on bone fragmentation, as it is a common taphonomic variable that effects many archaeofaunal assemblages across the globe. In particular, one of our investigations was to better understand the relationship between fragmentation and deposit type. To do this, we developed a Fragmentation Agency Index (FAI), which is defined as the ratio of percent unidentifiable (%Unidentifiable) to precent whole (%Whole) in an assemblage. %Whole measures extent of bone fragmentation by gauging the number of whole skeletal elements; it is a ratio of the number of whole specimens to the total number of specimens in an assemblage. %Unidentifiable coarsely measures intensity of fragmentation given the smaller fragments are the less identifiable criteria preserves (Cannon 2013). We calculate FAI by dividing fragmentation intensity (%Unidentifiable) by fragmentation extent (%Whole), which simplifies to: the number of unidentifiable specimens divided by the number of whole specimens. A higher FAI indicates a faunal assemblage dominated by more unidentifiable fragments relative to the number of whole specimens, which is what we view as a faunal assemblage with high fragmentation agency. We then calculated FAI for the faunal remains in every Study Unit—or unique area of deposition—in the Crow Canyon Research Database (Figure 17). This provides a way to diagnose deposits with particularly high levels of fragmentation in the central Mesa Verde region. The question now is, how do faunal remains (and their associated deposits) compare to other major sites in the region?
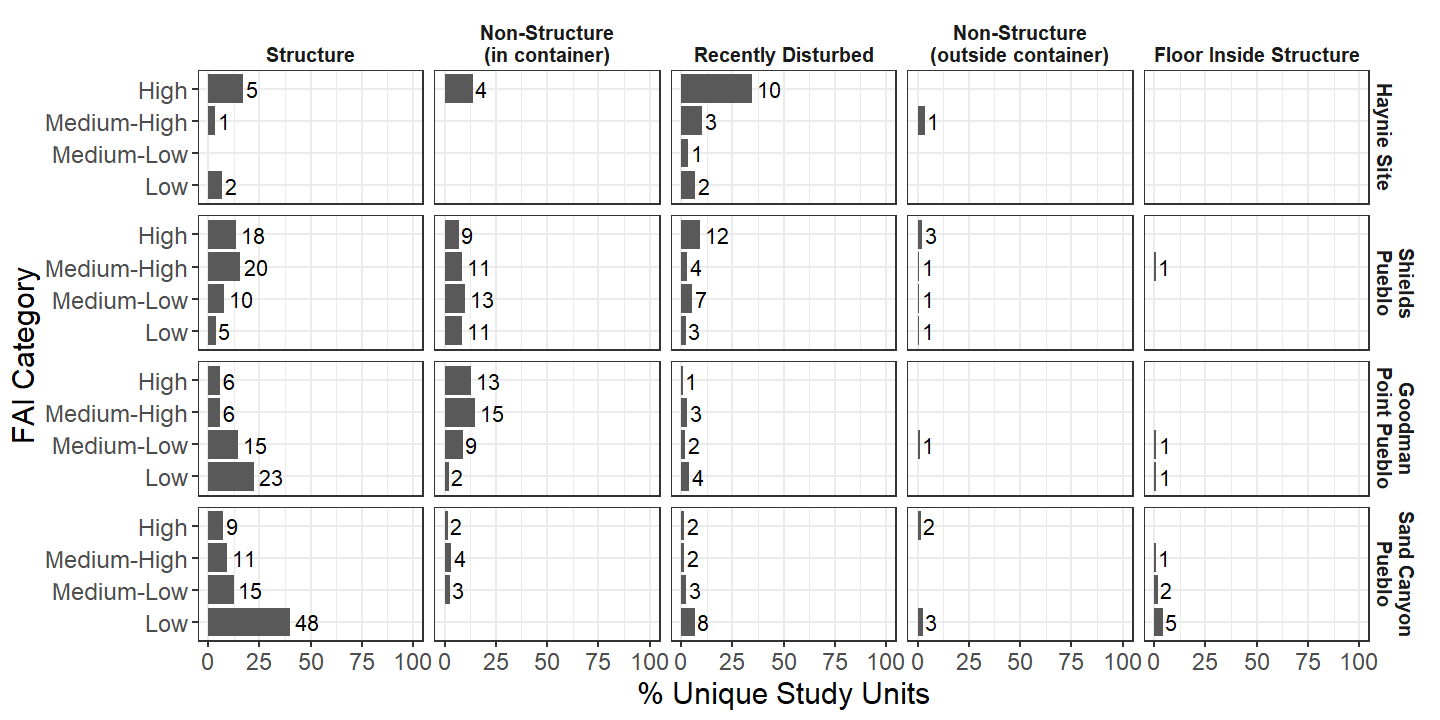
To compare FAI across major archaeological sites in the central Mesa Verde region, we broke down different FAI categories by deposits with different taphonomic potential. We created five different categories to capture taphonomic potential: primary deposits within a structure, secondary or other deposits shielded by a container of some kind (e.g., a structure), recently disturbed deposits (e.g., looter’s pits or chained areas), unshielded deposits (e.g., primary midden), or floor deposits. In that order, we term the deposits Structure, Non-Structure (in container), Recently Disturbed, Non-Structure (outside container), and Floor Inside Structure. We also put FAI into categories defined by quantile: below 25 is low, between 25 and 50 is medium-low, between 50 and 75 is medium-high, and higher than 75 is high. Using these grouping criteria, we see that Haynie is distinct from other major Crow Canyon projects in that the majority of faunal samples identified thus far come from Study Units that are recently disturbed with a high level of fragmentation agency (Figure 18). Shields Pueblo has a somewhat similar history of recent disturbance, but a much more even distribution of fragmentation agency across the FAI categories. Additionally, other Crow Canyon projects had a much higher focus of excavation in structures. The fragmentation of archaeofaunal remains identified from Haynie thus far are truly different compared to other sites in the Crow Canyon Research Database.
6 Conclusions
This year’s faunal report has added three new distinct areas of research associated with the Haynie site archaeofauna. First, we demonstrated—using stable isotope analysis and isoscape modeling—that the bison specimens recovered from the Haynie site do not come from the Plains region. It could be bison were procured closer to the Haynie site, and there are areas in the general Four Corners region that might be good candidates for areas of bison procurement. Second, preliminary evidence suggests mule deer may not have been as extensively overhunted at the end of Pueblo III period as previously thought. We have provided a snapshot of mule deer body size through time, of which Haynie importantly provided PII period samples. We have shown that signals of mule deer body size may not match with times where harvest pressure is thought to be intense. Future research will benefit from focusing on expanding astragali datasets, refining body size models specific to mule deer, and exploring alternative explanations associated with body size (i.e., environmental and/or ecological influences). Finally, to compare bone fragmentation across major archaeological sites in the central Mesa Verde region, we categorized deposits based on taphonomic potential and created an intuitive measure of fragmentation called the Fragmentation Agency Index (FAI). Haynie stands out in that most faunal samples from the site come from recently disturbed contexts with high fragmentation. This is different from other Crow Canyon projects, which typically emphasize excavation in structures and show more evenly distributed FAI patterns. While fragmentation is certainly distinctive at Haynie, additional research is needed to disentangle the cause of the high level of fragmentation.
7 Acknowledgements
Thank you to Jennifer Cotton for creating and sharing a new TIFF of her original isoscape for this project. Thanks as well to Blythe Morrison and Canyon of the Ancients Visitor Center and Museum for providing access to artiodactyl astragali used in this research.
8 References
Adams, Bradley J., and Pam J. Crabtree. 2012. Comparative Osteology a Laboratory and Field Guide of Common North American Animals. Boston, MA: Elsevier/Academic Press.
Andrews, Peter, and Jill Cook. 1990. Owls, Caves and Fossils: Predation, Preservation and Accumulation of Small Mammal Bones in Caves, with an Analysis of the Pleistocene Cave Faunas from Westbury-Sub-Mendip, Somerset, UK. Chicago, IL: University of Chicago Press.
Badenhorst, Shaw, and Jonathan C. Driver. 2009. “Faunal Changes in Farming Communities from Basketmaker II to Pueblo III (A.D. 1-1300) in the San Juan Basin of the American Southwest.” Journal of Archaeological Science 36 (9): 1832–41. https://doi.org/10.1016/j.jas.2009.04.006.
Badenhorst, Shaw, Jonathan C. Driver, and Steve Wolverton. 2023. “The Exploitation of Rodents in the Mesa Verde Region.” In Research, Education, and American Indian Partnerships at the Crow Canyon Archaeological Center, edited by Susan C. Ryan, 325–34. Salt Lake City: The University of Utah Press. https://doi.org/10.5876/9781646424597.c020.
Baker, Brenda J., Dupras Tosha L., and Matthew W. Tocheri. 2005. The Osteology of Infants and Children. Vol. 12. College Station, TX: Texas A&M University Press.
Baker, Bruce W., and Edward P. Hill. 2003. “Beaver (Castor Canadensis).” In Wild Mammals of North America: Biology, Management, and Conservation, edited by George A. Feldhamer, Bruce C. Thompson, and Joseph A. Chapman, 2nd ed., 288–310. Baltimore, MD: The Johns Hopkins University Press.
Beacham, E. Bradley, and Stephen R. Durand. 2007. “Eggshell and the Archaeological Record: New Insights into Turkey Husbandry in the American Southwest.” Journal of Archaeological Science 34 (10): 1610–21. https://doi.org/10.1016/j.jas.2006.11.015.
Beaglehole, Ernest. 1936. Hopi Hunting and Ritual. Yale University Publications in Anthropology 4. New Haven, CT: Yale University Press.
Bocinsky, R. Kyle, Jason A. Cowan, Timothy A. Kohler, and C. David Johnson. 2012. “How Hunting Changes the VEP World, and How the VEP World Changes Hunting.” In Emergence and Collapse of Early Villages: Models of Central Mesa Verde Archaeology, edited by Timothy A. Kohler and Mark Varien, 145–52. Berkeley: University of California Press.
Broughton, Jack M., Michael D. Cannon, Frank E. Bayham, and David A. Byers. 2011. “Prey Body Size and Ranking in Zooarchaeology: Theory, Empirical Evidence, and Applications from the Northern Great Basin.” American Antiquity 76 (3): 403–28. https://doi.org/10.7183/0002-7316.76.3.403.
Broughton, Jack M., and Shawn D. Miller. 2016. Zooarchaeology and Field Ecology: A Photographic Atlas. Salt Lake City: The University of Utah Press.
Cannon, Michael D. 2013. “NISP, Bone Fragmentation, and the Measurement of Taxonomic Abundance.” Journal of Archaeological Method and Theory 20 (3): 397–419. https://doi.org/10.1007/s10816-012-9166-z.
Chavez, Angela Marie. 2008. “Comparative Vertebral Morphology in Medium-Sized North American Artiodactyla.” The Artifact 46: 1–17.
Cloutier, Richard, and Peter L. Forey. 1991. “Diversity of Extinct and Living Actinistian Fishes (Sarcopterygii).” In The Biology of Latimeria Chalumnae and Evolution of Coelacanths, edited by John A. Musick, Michael N. Bruton, and Eugene K. Balon, 59–74. Springer.
Cohen, Alan, and D. Serjeantson. 1996. A Manual for the Identification of Bird Bones from Archaeological Sites. London: Archetype Publications.
Conrad, Cyler, Emily Lena Jones, Seth D. Newsome, and Douglas W. Schwartz. 2016. “One Isotopes, Eggshell and Turkey Husbandry at Arroyo Hondo Pueblo.” Journal of Archaeological Science: Reports 10: 566–74. https://doi.org/10.1016/j.jasrep.2016.06.016.
Cotton, Jennifer M., Thure E. Cerling, Kathryn A. Hoppe, Thomas M. Mosier, and Christopher J. Still. 2016. “Climate, CO2, and the History of North American Grasses Since the Last Glacial Maximum.” Science Advances 2 (3): e1501346. https://doi.org/10.1126/sciadv.1501346.
Dombrosky, Jonathan, Caitlin S. Ainsworth, Abigail A. Judkins, Jana Valesca Meyer, Michael A. Adler, and Emily Lena Jones. 2023. “Reconsidering Archaeological Garden Hunting: A View from the Northern U.S. Southwest.” Journal of Archaeological Science: Reports 51: 104194. https://doi.org/10.1016/j.jasrep.2023.104194.
Dombrosky, Jonathan, and Ahna Feldstein. 2024. “Year Two of Archaeofaunal Analysis at the Haynie Site (5MT1905).” In Excavation and Additional Studies at The Haynie Site (5MT1905) by the Crow Canyon Archaeological Center Annual Report 2023, edited by R. David Satterwhite, Susan C. Ryan, Kellam Throgmorton, Steve Copeland, Kate Hughes, Jamie Merewether, and Reuven Sinensky, 71–104. https://crowcanyon.org/wp-content/uploads/2024/02/NCOP-Annual-Report-2023_-Final.pdf.
Dombrosky, Jonathan, and Eric Gilmore. 2023. “Year One of Archaeofaunal Analysis at the Haynie Site (5MT1905).” In Excavation and Additional Studies at The Haynie Site (5MT1905) by the Crow Canyon Archaeological Center Annual Report 2022, edited by Kellam Throgmorton, Susan C. Ryan, Jonathan Dombrosky, Benjamin Bellorado, Steven Copeland, Kate Hughes, and Jamie Merewether, 45–88. https://www.crowcanyon.org/wp-content/uploads/2023/02/ncop-annual-report-2022-final.pdf.
Dombrosky, Jonathan, Thomas F. Turner, Alexandra Harris, and Emily Lena Jones. 2022. “Body Size from Unconventional Specimens: A 3D Geometric Morphometrics Approach to Fishes from Ancestral Pueblo Contexts.” Journal of Archaeological Science 142: 105600. https://doi.org/10.1016/j.jas.2022.105600.
Driver, Jonathan C. 1992. “Identification, Classification and Zooarchaeology.” Circaea 9 (1): 35–47.
———. 2002. “Faunal Variation and Change in the Northern San Juan Region.” In Seeking the Center Place: Archaeology and Ancient Communities in the Mesa Verde Region, edited by Mark D. Varien and Richard H. Wilshusen, 143–60. Salt Lake City: The University of Utah Press.
———. 2006. Crow Canyon Archaeological Center Manual for Description of Vertebrate Remains. 8th ed. Cortez, CO: Crow Canyon Archaeological Center.
———. 2011. “Human Impacts on Animal Populations in the American Southwest.” In Movement, Connectivity and Landscape Change in the Ancient Southwest, edited by Margaret C. Nelson and Colleen Strawhacker, 179–98. Boulder: University Press of Colorado.
———. 2011. “Identification, Classification and Zooarchaeology (Featured Reprint and Invited Comments).” Ethnobiology Letters 2: 19–39. https://doi.org/10.14237/ebl.2.2011.32.
Driver, Jonathan C., and Joshua R. Woiderski. 2008. “Interpretation of the "Lagomorph Index" in the American Southwest.” Quaternary International 185: 3–11. https://doi.org/10.1016/j.quaint.2007.09.022.
Elbroch, Mark. 2006. Animal Skulls: A Guide to North American Species. Mechanicsburg, PA: Stackpole Books.
Ellyson, Laura. 2014. “Resource Intensification of Small Game Use at Goodman Point, Southwestern Colorado.” Master's Thesis, Denton: Department of Geography, University of North Texas. https://digital.library.unt.edu/ark:/67531/metadc699883/.
Ellyson, Laura J., Lisa Nagaoka, and Steve Wolverton. 1990. “Animal Resource Use Related to Socioenvironmental Change Among Mesa Verde Farmers.” Journal of Anthropological Research 75 (3): 361–92. https://doi.org/10.1086/704143.
Fernández-Jalvo, Yolanda, and Peter Andrews. 2016. Atlas of Taphonomic Identifications: 1001+ Images of Fossil and Recent Mammal Bone Modification. Springer.
France, Diane L. 2009. Human and Nonhuman Bone Identification: A Color Atlas. Boca Raton, FL: CRC Press.
Gilbert, B. Miles. 1980. Mammalian Osteology. Laramie, WY: B. M. Gilbert.
Gilbert, B. Miles, Larry D. Martin, and Howard G. Savage. 1981. Avian Osteology. Laramie, WY: B. M. Gilbert.
Gobalet, Kenneth W. 2001. “A Critique of Faunal Analysis: Inconsistency Among Experts in Blind Tests.” Journal of Archaeological Science 28 (4): 377–86. https://doi.org/10.1006/jasc.2000.0564.
Grayson, Donald K. 1984. Quantitative Zooarchaeology: Topics in the Analysis of Archaeological Faunas. Orlando, FL: Academic Press. https://www.sciencedirect.com/book/9780122972805/quantitative-zooarchaeology.
Guiry, Eric, Trevor J. Orchard, Suzanne Needs-Howarth, and Paul Szpak. 2021. “Isotopic Evidence for Garden Hunting and Resource Depression in the Late Woodland of Northeastern North America.” American Antiquity 86 (1): 90–110. https://doi.org/10.1017/aaq.2020.86.
Hargrave, Lyndon L. 1965. “Archaeological Bird Bones from Chapin Mesa, Mesa Verde National Park.” American Antiquity 31 (2): 156–60. https://doi.org/10.1017/S0081130000004494.
Hargrave, Lyndon L., and Steven D. Emslie. 1979. “Osteological Identification of Sandhill Crane Versus Turkey.” American Antiquity 44 (2): 295–99. https://doi.org/10.2307/279079.
Hillson, Simon. 1986. Teeth. New York: Cambridge University Press.
———. 1996. Mammal Bones and Teeth: An Introductory Guide to Methods of Identification. London: Institute of Archaeology, University College London.
Jacobson, Jodi A. 2003. “Identification of Mule Deer (Odocoileus Hemionus) and White-Tailed Deer (Odocoileus Virginianus) Postcranial Remains as a Means of Determining Human Subsistence Strategies.” Plains Anthropologist 48 (187): 287–97. https://doi.org/10.1080/2052546.2003.11949269.
Ladd, Edmund J. 1963. “Zuni Ethno-Ornithology.” Master's Thesis, Albuquerque: Department of Anthropology, University of New Mexico.
Lapham, Heather A., Gary M. Feinman, and Linda M. Nicholas. 2016. “Turkey Husbandry and Use in Oaxaca, Mexico: A Contextual Study of Turkey Remains and SEM Analysis of Eggshell from the Mitla Fortress.” Journal of Archaeological Science: Reports 10: 534–46. https://doi.org/10.1016/j.jasrep.2016.05.058.
Lawrence, Barbara. 1951. Post-Cranial Skeletal Characters of Deer, Pronghorn, and Sheep-Goat, with Notes on Bos and Bison. Papers of the Peabody Museum of American Archaeology and Ethnology. Cambridge, MA.
LaZar, Miranda, and Jonathan Dombrosky. 2022. “Exploring Raptor Exchange.” Archaeology Southwest Magazine 35 (1 & 2): 29–31. https://www.archaeologysouthwest.org/product/asw35-1-2/.
Lyman, R. Lee. 1994. Vertebrate Taphonomy. Cambridge: Cambridge University Press.
———. 2002. “Taxonomic Identification of Zooarchaeological Remains.” The Review of Archaeology 23 (2): 13–20.
———. 2008. Quantitative Paleozoology. Cambridge: Cambridge University Press.
———. 2019. “Assumptions and Protocol of the Taxonomic Identification of Faunal Remains in Zooarchaeology: A North American Perspective.” Journal of Archaeological Method and Theory 26 (4): 1376–1438. https://doi.org/10.1007/s10816-019-09414-0.
Martin, Jeff M., Rachel A. Martin, and Jim I. Mead. 2017. “Late Pleistocene and Holocene Bison of the Colorado Plateau.” The Southwestern Naturalist 62 (1): 14–28. https://doi.org/10.1894/0038-4909-62.1.14.
Minckley, Wendell L., and Paul C. Marsh. 2009. Inland Fishes of the Greater Southwest: Chronicle of a Vanishing Biota. Tucson: The University of Arizona Press.
Monagle, Victoria, and Emily Lena Jones. 2020. “Dog Life and Death in an Ancestral Pueblo Landscape.” In Dogs: Archaeology beyond Domestication, edited by Brandi Bethke and Amanda Burtt, 45–71. Gainesville: University Press of Florida.
Muir, Robert J., and Jonathan C. Driver. 2002. “Scale of Analysis and Zooarchaeological Interpretation: Pueblo III Faunal Variation in the Northern San Juan Region.” Journal of Anthropological Archaeology 21 (2): 165–99. https://doi.org/10.1006/jaar.2001.0392.
Munro, Natalie D. 2011. “Domestication of the Turkey in the American Southwest.” In The Subsistence Economies of Indigenous North American Societies: A Handbook, edited by Bruce D. Smith, 543–55. Washington D.C.: Smithsonian Institution Scholarly Press.
Nelson, Joseph S. 2006. Fishes of the World. Hoboken, NJ: John Wiley.
Olsen, Stanley John. 1964. Fish, Amphibian and Reptile Remains from Archaeological Sites. Cambridge, MA: Peabody Museum.
———. 1964. Mammal Remains from Archaeological Sites. Cambridge, MA: Peabody Museum.
———. 1979. Osteology for the Archaeologist. Cambridge, MA: Peabody Museum.
Purdue, James R. 1987. “Estimation of Body Weight of White-Tailed Deer (Odocoileus Virginianus) from Bone Size.” Journal of Ethnobiology 7 (1): 1–12. https://ethnobiology.org/sites/default/files/pdfs/JoE/7-1/Purdue1987.pdf.
———. 1989. “Changes During the Holocene in the Size of White-Tailed Deer (Odocoileus Virginianus) from Central Illinois.” Quaternary Research 32 (3): 307–16. https://doi.org/10.1016/0033-5894(89)90097-5.
R Core Team. 2024. R: A Language and Environment for Statistical Computing. Vienna, Austria: R Foundation for Statistical Computing. https://www.R-project.org/.
Rawlings, Tiffany A. 2006. “Faunal Analysis and Meat Procurement: Reconstructing the Sexual Division of Labor at Shields Pueblo, Colorado.” Doctoral Dissertation, Burnaby, BC: Department of Archaeology, Simon Fraser University. https://summit.sfu.ca/item/2374.
Roze, Uldis, and Linda M. Ilse. 2003. “Porcupine (Erethizon Dorsatum).” In Wild Mammals of North America: Biology, Management, and Conservation, edited by George A. Feldhamer, Bruce C. Thompson, and Joseph A. Chapman, 2nd ed., 371–80. Baltimore, MD: The Johns Hopkins University Press.
Ryan, Susan C. 2016. “The Northern Chaco Outliers Project: A Proposal to Conduct Archaeological Testing at the Haynie Site, Southwestern Colorado.” Manuscript on file Crow Canyon Archaeological Center, Cortez, CO. Submitted to Colorado State Historic Preservation Office, Denver, CO.
Schollmeyer, Karen Gust, and Jonathan C. Driver. 2023. “Fine-Grained Chronology Reveals Human Impacts on Animal Populations in the Mesa Verde Region of the American Southwest.” In Research, Education and American Indian Partnerships at the Crow Canyon Archaeological Center, edited by Susan C. Ryan, 335–46. Boulder: University of Colorado Press.
Smart, Tamela S. 2009. “Carpals and Tarsals of Mule Deer, Black Bear and Human: An Osteology Guide for the Archaeologist.” Master's Thesis, Pullman: Department of Anthropology, Western Washington University. https://cedar.wwu.edu/cgi/viewcontent.cgi?referer=&httpsredir=1&article=1018&context=wwuet.
Speth, John D. 1991. “Some Unexplored Aspects of Mutualistic Plains-Pueblo Food Exchange.” In Farmers, Hunters, and Colonists: Interaction Between the Southwest and the Southern Plains, 18–35. University of Arizona Press. http://www.jstor.org/stable/j.ctvss3wqx.5.
Spielmann, Katherine A., and Eric A. Angstadt-Leto. 1996. “Hunting, Gathering, and Health in the Prehistoric Southwest.” In Evolving Complexity and Environmental Risk in the Prehistoric Southwest, edited by Joseph A. Tainter, and Bonnie B. Tainter, 79–106. Reading, MA: CRC Press.
Staudenmaier, Anna R, Lisa A. Shipley, Andris J. Bibelnieks, Meghan J. Camp, and Daniel H. Thornton. 2021. “Habitat Use and Spatio-Temporal Interactions of Mule and White-Tailed Deer in an Area of Sympatry in NE Washington.” Ecosphere 12 (11): e03813. https://doi.org/10.1002/ecs2.3813.
Sublette, James E., Michael D. Hatch, and Mary F. Sublette. 1990. The Fishes of New Mexico. Albuquerque: University of New Mexico Press.
Throgmorton, Kellam, Susan C. Ryan, Benjamin Bellorado, Jeremy Grundvig, Steve Copeland, and Timothy Wilcox. 2022. “Excavation and Additional Studies at The Haynie Site (5MT1905) by the Crow Canyon Archaeological Center.” Annual Report 2021. Cortez, CO: Crow Canyon Archaeological Center. https://www.crowcanyon.org/wp-content/uploads/2022/02/ncop-annual-report-2021-final-1.pdf.
Throgmorton, Kellam, Susan C. Ryan, Jonathan Dombrosky, Benjamin Bellorado, Steven Copeland, Kate Hughes, and Jamie Merewether. 2023. “Excavation and Additional Studies at The Haynie Site (5MT1905) by the Crow Canyon Archaeological Center.” Annual Report 2022. Cortez, CO: Crow Canyon Archaeological Center. https://crowcanyon.org/wp-content/uploads/2023/02/ncop-annual-report-2022-final.pdf.
Tyler, Hamilton A. 1991. Pueblo Birds and Myths. Flagstaff, AZ: Northland Publishing Company.
Weitzel, Elic M. 2023. “Resilience of White-Tailed Deer (Odocoileus virginianus) to Human Hunting in Precolonial New England: The Faunal Remains from the Morgan Site (6HT120), Rocky Hill, Connecticut.” Journal of Archaeological Science: Reports 41: 103913. https://doi.org/10.1016/j.jasrep.2023.103913.
White, Tim D., Michael T. Black, and Pieter A. Folkens. 2012. Human Osteology. 3rd ed. Cambridge, MA: Academic Press.
Wickham, Hadley. 2010. “A Layered Grammar of Graphics.” Journal of Computational and Graphical Statistics 19 (1): 3–28. https://doi.org/10.1198/jcgs.2009.07098.
Wickham, Hadley, Mara Averick, Jennifer Bryan, Winston Chang, Lucy D’Agostino McGowan, Romain François, Garrett Grolemund, Alex Hayes, Lionel Henry, and Jim Hester. 2019. “Welcome to the Tidyverse.” Journal of Open Source Software 4 (43): 1686. https://doi.org/10.21105/joss.01686.
Wolverton, Steve. 2008. “Harvest Pressure and Environmental Carrying Capacity: An Ordinal-Scale Model of Effects on Ungulate Prey.” American Antiquity 73 (2): 179–99. https://doi.org/10.1017/S0002731600042244.
———. 2013. “Data Quality in Zooarchaeological Faunal Identification.” Journal of Archaeological Method and Theory 20 (3): 381–96. https://doi.org/10.1007/s10816-012-9161-4.
Wolverton, Steve, James H. Kennedy, and John D. Cornelius. 2007. “A Paleozoological Perspective on White-Tailed Deer (Odocoileus Virginianus Texana) Population Density and Body Size in Central Texas.” Environmental Management 39 (4): 545–52. https://doi.org/10.1007/s00267-006-0240-8.
Wolverton, Steve, Lisa Nagaoka, Julie Densmore, and Ben Fullerton. 2008. “White-Tailed Deer Harvest Pressure & Within-Bone Nutrient Exploitation During the Mid-to Late Holocene in Southeast Texas.” Before Farming 2: 1–23. https://doi.org/10.3828/bfarm.2008.2.3.
Wolverton, Steve, Michael A. Huston, James H. Kennedy, Kevin Cagle, and John D. Cornelius. 2009. “Conformation to Bergmann’s Rule in White-Tailed Deer Can Be Explained by Food Availability.” American Midland Naturalist 162 (2): 403–17. https://doi.org/10.1674/0003-0031-162.2.403.
Wolverton, Steve, and Lisa Nagaoka. 2018. “Zooarchaeology: Investigating Past Interactions Between Humans and Other Animals.” In Ethnozoology, edited by Rômulo Romeu Nóbrega Alves and Ulysses Paulino Albuquerque, 25–43. Cambridge, MA: Academic Press. https://doi.org/10.1016/B978-0-12-809913-1.00003-X.
Citation
BibTeX citation:
@incollection{dombrosky2025,
author = {Dombrosky, Jonathan and Betts, Chelsea and Wolverton, Steve
and C. Ryan, Susan},
editor = {David Satterwhite, R. and C. Ryan, Susan and Copeland, Steve
and Sinensky, Reuven and Merewether, Jamie and Hughes, Kate},
title = {Year {Three} of {Archaeofaunal} {Analysis} at the {Haynie}
{Site} {(5MT1905)}},
booktitle = {Excavation and Additional Studies at The Haynie Site
(5MT1905) by the Crow Canyon Archaeological Center Annual Progress
Report 2024},
pages = {45-79},
date = {2025},
url = {https://crowcanyon.org/wp-content/uploads/2025/02/Annual-Report-2024_-NoOsteo.pdf},
langid = {en}
}
For attribution, please cite this work as:
Dombrosky, Jonathan, Chelsea Betts, Steve Wolverton, and Susan C. Ryan.
2025. “Year Three of Archaeofaunal Analysis at the Haynie Site
(5MT1905).” In Excavation and Additional Studies at The
Haynie Site (5MT1905) by the Crow Canyon Archaeological Center Annual
Progress Report 2024, edited by R. David Satterwhite, Susan C.
Ryan, Steve Copeland, Reuven Sinensky, Jamie Merewether, and Kate
Hughes, 45–79. https://crowcanyon.org/wp-content/uploads/2025/02/Annual-Report-2024_-NoOsteo.pdf.